光合成の測定
植物の生育と光合成
アリストテレスの時代には、植物は、いわば「自然に」大きくなると考えられていました。動物を見ていれば、いかにも食事をすることによって大きくなっているように見える上に、自分たち人間も、ものを食べないでいれば力が出なくなることを経験しています。それにひきかえ、植物は、動くでもなく、何かを体に取り入れているようにも見えないのに、いつの間にか大きくなっていきます。古代の人が、植物が自然に大きくなると考えたのも不思議はありません。
ようやく1600年代の半ばになってVan Helmontが有名な柳の木の実験で、植物の生育にとって水が重要であることを見いだしました。Van Helmont自身は、科学者と錬金術師の中間に位置するような人物で、彼の実験も、元々はギリシャの哲学者ターレスの「万物は水である」という哲学を実証するために行なわれたものでした。物質の出入りに関して、ガス(酸素、二酸化炭素)の存在に気がついていなかったためにVan Helmontは「植物は水からできている」という誤った結論を得ましたが、この実験は、結論の妥当性とは別に、生物の実験に定量性をもたらしたという点で極めて重要です。
その後、Mariotte やHalesが、植物が二酸化炭素を吸収していることを見いだし、また、Priestleyが酸素を発生していることを見いだしました。さらに、Ingen-Housz とSenebierが1700年代の終わりに、植物の酸素発生には光が必要であることを明らかにして、光合成の基本式
二酸化炭素+水+光 → 光合成産物+酸素
が完成しました。その後、光合成の研究は進み、現在までに10人以上のノーベル賞受賞者を輩出しながら数多くの研究者によってその構造・機能の詳細が明らかになってきました。光合成は植物の生育を通して生物界全体のエネルギー生産を担っているため、現在も、基礎研究はもちろん、農業生産や地球環境など様々な立場から研究が盛んに行われています。
光合成の測定方法
植物は、その生存に必要なエネルギーを光合成に依存しているため、植物の生育を考えるにあたって光合成の速度を測ることは極めて重要です。光合成の速度を測るためには、上記の光合成の基本式の右辺でも左辺でもどれかの物質の量の変化を測定すればよいことになります。この中で、水は、植物体がもともとかなりの量を持っている上に、根からの吸水と葉からの蒸散によって常に入れ替わっているため、光合成の測定に使うことは不可能です。何しろ、蒸散が盛んな葉では、葉の中の水が1時間で4回入れ替わる計算になり、光合成の基質として使われる水は、蒸散量のたった0.4%といわれています。そのような状況で水の量を測定しても光合成に関する情報を得ることはできません。
とすると、残りは、二酸化炭素、酸素、光合成産物です。また特殊な条件下では光を使って光合成を測定することも可能です。以下に、それらの様々な光合成の測定方法を見ていきたいと思います。
光合成産物量の測定
(1)ヨウ素デンプン反応
光合成の基本式を考えれば、単純に光合成産物量の増加を測定することにより光合成の速度を見積もることができるはずです。学校の実験などでおなじみの、ヨウ素デンプン反応によるデンプンの検出はこれを利用しています。ヨウ素デンプン反応は、定量性には欠けるものの、手軽に実験できるために広く使われています。しかし、問題点として、全ての植物がデンプンを蓄積するわけではないということがあります。
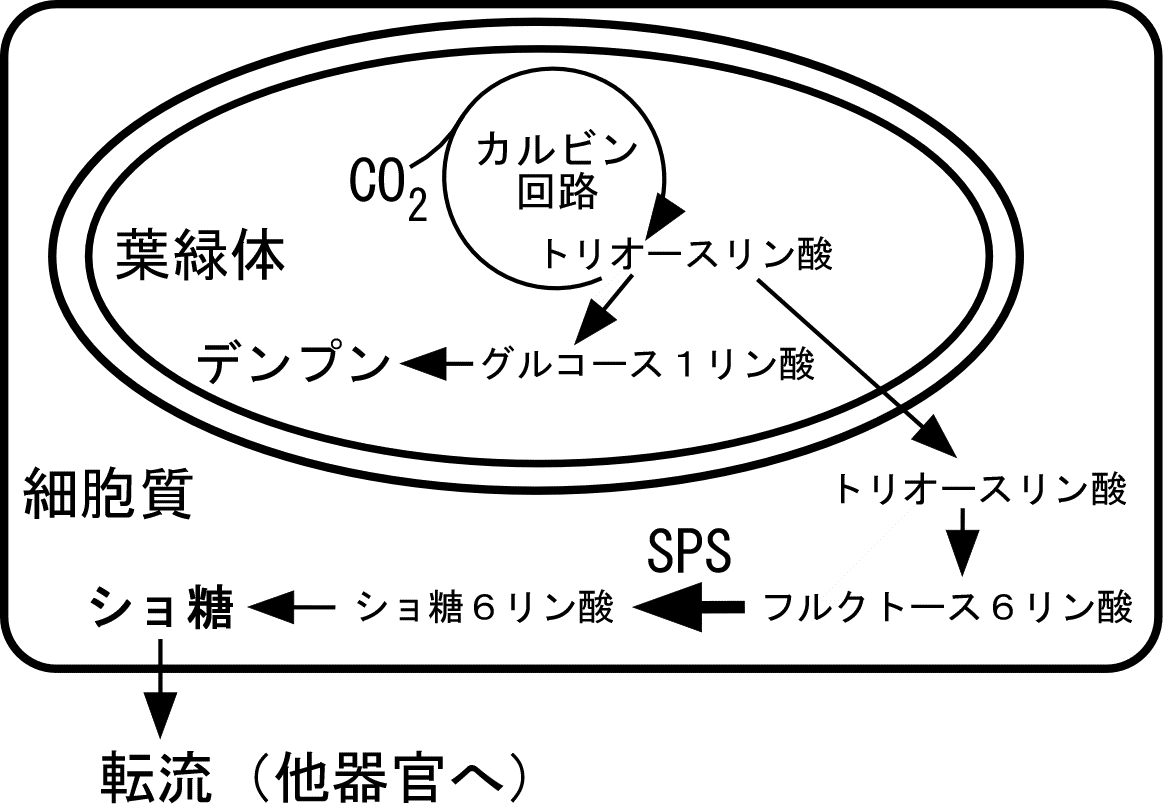 右の図のように、葉におけるデンプンは葉緑体内で光合成産物から合成されますが、一方で、光合成産物は葉緑体からトリオースリン酸の形で細胞質に輸送され、細胞質でショ糖(植物種によってはソルビトールなどの場合もある)になった後、転流によって他の器官に運ばれます。ショ糖の合成は、細胞質のショ糖リン酸合成酵素(Sucrose Phosphate Synthase、SPSと略称される)によるショ糖6リン酸の合成が律速段階となっています。SPSの活性の低い場合には転流の速度は低く、葉緑体にはデンプンが蓄積します。一方、SPSの活性が高いと、転流速度は高く、葉緑体にはあまりデンプンが蓄積しません。植物によってSPSの活性は異なり、また、葉緑体にどの程度デンプンを蓄積するかも異なります。例えば、イネ科の植物の場合は葉緑体にはほとんどデンプンを蓄積しないものがおおくあります。従って、ヨウ素デンプン反応による光合成産物の検出は、そのような植物には用いることができない点に注意する必要があります。ヨウ素デンプン反応に関しては「ヨウ素デンプン反応の実際」のページに実験例を載せておきました。
右の図のように、葉におけるデンプンは葉緑体内で光合成産物から合成されますが、一方で、光合成産物は葉緑体からトリオースリン酸の形で細胞質に輸送され、細胞質でショ糖(植物種によってはソルビトールなどの場合もある)になった後、転流によって他の器官に運ばれます。ショ糖の合成は、細胞質のショ糖リン酸合成酵素(Sucrose Phosphate Synthase、SPSと略称される)によるショ糖6リン酸の合成が律速段階となっています。SPSの活性の低い場合には転流の速度は低く、葉緑体にはデンプンが蓄積します。一方、SPSの活性が高いと、転流速度は高く、葉緑体にはあまりデンプンが蓄積しません。植物によってSPSの活性は異なり、また、葉緑体にどの程度デンプンを蓄積するかも異なります。例えば、イネ科の植物の場合は葉緑体にはほとんどデンプンを蓄積しないものがおおくあります。従って、ヨウ素デンプン反応による光合成産物の検出は、そのような植物には用いることができない点に注意する必要があります。ヨウ素デンプン反応に関しては「ヨウ素デンプン反応の実際」のページに実験例を載せておきました。
(2)半葉法(葉半法)
古くは、光合成産物の増加に特定せず、植物の葉の重量全体の増加を測定することによって光合成活性を見積もることが行なわれました。半葉法というのは、光合成の同化産物がデンプンであることを見いだしたSachsが1883年に考案した方法で、1枚の葉の半分にだけ光をあて、光をあてた側と光をあてなかった側から同じ面積の葉を切り出して、その乾燥重量の差を光合成の同化産物の蓄積によると考える方法です。
しかし、上記のように同化産物はいつまでも葉の中にとどまるわけではなく、転流によって他の器官に運ばれるので、葉の重量の増加は光合成の同化産物の蓄積よりも小さくなってしまいます。これを避けるために、野本と佐伯によって熱ガードリング法という改良法が1969年に提案されました。これは、測定前に葉柄を高温の水蒸気により処理し、師部をいわば「殺す」ことによって転流を抑えるものです。これにより、転流による光合成産物の減少は抑えられますが、明確な差を得ようと光合成させる時間を増やすと別な問題が生じます。
一般に、生体内の代謝系はフィードバック制御がかかっており、ある反応が進みすぎた場合は、その反応を抑えるようなメカニズムが働きます。光合成の場合も同じで、光合成産物が蓄積すると光合成関連遺伝子の発現が抑えられ、光合成活性が低下します。従って、転流を阻害した場合には、光合成産物が蓄積することによって、光合成活性が本来のレベルより低くなってしまいます。このような副作用は、時間を短くすれば無視できるようになりますが、その場合には、光合成産物量の増加も少ないので、測定誤差は大きくなってしまいます。このような問題点もあって、現在ではこの方法は光合成の研究には使われていません。
酸素発生の測定
(1)検圧法
1910年代にWarburgはガスの発生量により光合成活性を測定する検圧法を開発しました。液相での光合成を考えた場合、光合成により発生する酸素の水への溶解度は高くないので、ある一定以上の量になると気体になります。つまり、液相に溶けている二酸化炭素を基質として使っている限り、光合成による酸素の発生は、気体の発生量として測定し得ることになります。気相の場合にも、緩衝液を使って二酸化炭素濃度を一定に保てば、この方法を使うことができます。検圧法は、現在の研究レベルからすると精度が低いこと、また、温度など他の条件の影響を受けやすいことから、実際の研究現場では使われなくなっています。しかし、同様の原理のプロダクトメーターは、酸素の発生が視覚的に把握できること、装置の値段が数万円と比較的安いことから、現在でも教育現場では使われています。また、水を詰めたペットボトルとオオカナダモなどの水草を用いる酸素発生の確認方法も、この検圧計による光合成測定のバリエーションとして考えることができます。
(2)明暗瓶法
この方法は、酸素瓶と呼ばれる密閉容器にプランクトンを含む水などの試料を入れ、1つを暗所、1つを明所において、一定時間の後、瓶の中の溶存酸素濃度を定量して、その差を光合成による酸素発生の活性の目安とするものです。溶存酸素の定量には、一般にウィンクラー法というヨウ素を用いた滴定法が使われます。海洋や湖沼に酸素瓶を沈めれば、現場の光環境・温度での光合成を測定できるので、水圏生態学の分野では広く用いられてきました。
この方法では、短い時間変化を追うことができないため、光合成のメカニズムの研究には用いられず、また、生態学的な研究の分野においても、後述するクロロフィル蛍光を用いた測定に取って代わられてきています。
(3)酸素電極
酸素電極は、陰極に白金、陽極に銀を用いた電極で、電極部に飽和塩化カリウム溶液を満たして電極間に電圧を印加すると、陰極では酸素の還元
O2 + 4H+ + 4e- → 2H2O
陽極では塩素の酸化
4Ag + 4Cl- → 4AgCl + 4e-
が起こって電流が流れます。この電流は溶液中の酸素濃度に比例するので、電流変化を測定することにより溶液中の変化を知ることができます。
 酸素電極の陰極は他の陽イオンとも反応し電流を流してしまうため、溶液中で直接反応を行うと液中のイオンの影響を受け、測定が不安定になります。通常用いられる、右の写真のようなクラーク型電極では、このような影響を除去するため、酸素は通過させるがイオンを通さないテフロン膜で電極を覆うことにより電極液と測定試料を分離し、測定試料の酸素濃度と平衡状態にある電極液の酸素濃度を測定します。酸素電極による測定は、温度の影響を強く受けるので、写真のように測定液を入れる容器の周りにさらに一定温度の水を循環させています。右から出ている光ファイバーは、光合成を行なわせるための光を照射するためのものです。通常、溶液中の酸素濃度の絶対値に対して短い時間の酸素濃度の変化率は小さいので、得られた信号から一定の大きさを引き算して(これをバッキングといいます)、結果を増幅して記録計などに出力します。
酸素電極の陰極は他の陽イオンとも反応し電流を流してしまうため、溶液中で直接反応を行うと液中のイオンの影響を受け、測定が不安定になります。通常用いられる、右の写真のようなクラーク型電極では、このような影響を除去するため、酸素は通過させるがイオンを通さないテフロン膜で電極を覆うことにより電極液と測定試料を分離し、測定試料の酸素濃度と平衡状態にある電極液の酸素濃度を測定します。酸素電極による測定は、温度の影響を強く受けるので、写真のように測定液を入れる容器の周りにさらに一定温度の水を循環させています。右から出ている光ファイバーは、光合成を行なわせるための光を照射するためのものです。通常、溶液中の酸素濃度の絶対値に対して短い時間の酸素濃度の変化率は小さいので、得られた信号から一定の大きさを引き算して(これをバッキングといいます)、結果を増幅して記録計などに出力します。
この方法では、秒単位で光合成速度の変化を調べることができるため、光合成メカニズムの研究に広く使われてきました。感度もよく、葉緑体やチラコイド膜の光合成の測定には欠かせない装置です。また、阻害剤などを用いると、光化学系Iや光化学系IIなどの部分反応の活性も測定できる利点があります。葉での測定などのために、気相の酸素電極も存在しますが、空気中の酸素濃度は21%にも達するため、その中で酸素濃度の小さな変化を測定するためには困難も多く、広く使われるには至っていません。
液相の酸素電極は、電極自体は数万円で購入できますが、上述のように、バッキング回路を手作りするか、バッキング回路付きの記録計を購入する必要があることと、慣れない人にはテフロン膜の扱いが難しいという欠点があります。
二酸化炭素吸収の測定
一般に、1原子分子のガスおよび単一の元素からなる2原子分子ガスは、赤外領域に吸収を持ちませんが、非対称な分子のガスは、赤外領域に吸収を持ちます。従って、空気の主成分である、窒素、酸素、アルゴン、二酸化炭素、水蒸気のうち、二酸化炭素と水蒸気だけが赤外領域の吸収を持ちます。光合成による二酸化炭素の吸収を測定するには、赤外領域の吸収の減少を測定すればよいことになりますが、その具体的な方法は複数考えられます。装置としては、ある程度の性能のものだと数百万円になるので、どうしても研究目的に限定されます。以下に、測定の具体的な仕組みを簡単に述べます。
(1)試料室の仕組み
 密閉容器中に葉を入れて光を当てれば、光合成によって容器内の二酸化炭素濃度は徐々に減少します。この減少速度を見ることによって、光合成速度を見積もる方法が、closed systemです。この場合、基質となる二酸化炭素濃度が徐々に減少するため、光合成速度も低下していくため、定常状態の測定ができないという欠点がありますが、閉鎖系なので、光合成速度がかなり低い場合でも測定が可能です。この方法の場合、二酸化炭素濃度変化の微分値をその時の二酸化炭素濃度に対してプロットすれば、一回の測定で光合成速度の二酸化炭素濃度依存性(A-Ciカーブ:AはAssimilation同化速度、Ciは葉内二酸化炭素濃度)を測定できます。
密閉容器中に葉を入れて光を当てれば、光合成によって容器内の二酸化炭素濃度は徐々に減少します。この減少速度を見ることによって、光合成速度を見積もる方法が、closed systemです。この場合、基質となる二酸化炭素濃度が徐々に減少するため、光合成速度も低下していくため、定常状態の測定ができないという欠点がありますが、閉鎖系なので、光合成速度がかなり低い場合でも測定が可能です。この方法の場合、二酸化炭素濃度変化の微分値をその時の二酸化炭素濃度に対してプロットすれば、一回の測定で光合成速度の二酸化炭素濃度依存性(A-Ciカーブ:AはAssimilation同化速度、Ciは葉内二酸化炭素濃度)を測定できます。
一方で、葉をおいた容器に一定濃度の二酸化炭素を含んだ空気を通気し、容器から出てくる空気の二酸化炭素濃度の減少によって光合成速度を見積もる方法が、open systemです。定常状態の光合成の測定には、通常こちらの方法が使われます。右の写真は、このopen systemの測定装置の例で、右側に葉を上下からはさむような形で試料室があり、左側が、制御部分です。
(2)水蒸気の影響の除去
上記のように、二酸化炭素の他に水蒸気も赤外に吸収を持つので、その影響を排除する必要があります。そのためには、(A)乾燥剤などで、物理的に水蒸気を取り除く、(B)湿度を測定して、水の分の吸収を補正する、(C)二酸化炭素と水蒸気で異なる吸収を持つ波長領域2カ所以上で赤外吸収を測定することにより、二酸化炭素、水蒸気、それぞれの赤外吸収への寄与を計算する、という3つの方法が考えられます。これらの方式には一長一短があり、測定装置のメーカーによって使う方式は異なります。
(3)葉内二酸化炭素濃度の計算
葉における光合成は、葉の中の二酸化炭素濃度に依存しますが、これは気孔の開度によって大きく影響を受けます。従って、測定容器の中の二酸化炭素濃度を一定にしても、葉の中の二酸化炭素濃度や光合成速度は、気孔の状態によって変わってしまいます。これを避けるために、通常、光合成速度は、葉の中の二酸化炭素濃度(Ci)をそろえて比較します。それでは、葉内二酸化炭素濃度はどのようにして測定できるのでしょうか。
気孔を二酸化炭素が出入りする速度(=光合成速度P)は、1)気孔の開度(ガスの通しやすさ:α)、2)外の二酸化炭素濃度(Ca)と葉内二酸化炭素濃度(Ci)の差、そして3)二酸化炭素の拡散しやすさ(ガスの通しやすさのガスの種類による違い:kC)、の3つによって決まります。
P=αkC(Ca-Ci)
気孔は、二酸化炭素の取り込み口であるとともに、水の蒸散のための排出口でもあります。従って水蒸気についても、気孔の開度、外の水蒸気濃度(Ha)と葉の中の水蒸気濃度(Hi)の差、水蒸気の拡散しやすさ(kH)の3つで蒸散速度(E)が決まります。
E=αkH(Hi-Ha)
ここで、光合成速度、蒸散速度は、実際にどれだけ二酸化炭素濃度が減少したか、水蒸気が増えたか、によって見積もることができ、また、外側の二酸化炭素濃度と水蒸気濃度も実測できます。さらに、葉の中の水蒸気濃度は飽和水蒸気濃度であると考えられ、また、二酸化炭素と水蒸気の拡散のしやすさは実測されています。従って、二酸化炭素と水蒸気に関する2つの式の中で、未知数は気孔開度α(2つの式に共通)と葉内の二酸化炭素濃度Ciの2つだけとなるので、連立方程式を解けば、葉内二酸化炭素濃度を見積もることができます。
「光」の測定
近年、光を用いて光合成を測定する方法が急速に発達しました。光合成の基本式
二酸化炭素+水+光 → 光合成産物+酸素
をエネルギーの面から見ると
照射光 → 生物的エネルギー+熱+蛍光
となります。これは、当てた光のエネルギーの一部は、光合成の結果、光合成産物などに生物的エネルギーとして蓄えられ、残りのエネルギーは熱もしくは蛍光として失われることを意味します。蛍光というのは、蛍光灯の蛍光と同じで、光エネルギーを吸収した色素が、そのエネルギーの一部を光として再放出した時にその光を蛍光といいます。エネルギーの面から見た上記の式を見ると、照射光を一定にしておいた場合に、もし熱になる割合が一定であれば、光合成が盛んであれば、クロロフィルから出る蛍光は小さく、光合成が止まれば、クロロフィル蛍光は大きくなることが予想されます。つまり、一定の光を葉に当てて、出てくる蛍光を測定することにより、光合成の状態をモニターすることができることになります。
実際には、照射光の種類や強度を様々に工夫し、また、測定する光を変調する(連続光ではなく、周期的な短いパルス光を使うことを意味します−AMラジオの原理と同じ)ことにより、光合成に関する様々な情報を得ることができます。原理と得られる情報の詳細は、光合成とクロロフィル蛍光をご覧ください。現在、様々なタイプの測定装置が販売されていますが、一番簡便なタイプの装置でも100万円程度するので、これも、ほぼ研究目的に限られてしまっています。以下では、実際の測定例を見てみましょう。
(1)パルス変調クロロフィル蛍光測定
 パルス変調という手法の導入により、クロロフィル蛍光の測定は、極めて高感度に行なえるようになりました。その結果、右の写真のように、植物の葉に測定装置のグラスファイバーの先端を近づけるだけで光合成の測定が可能になりました。測定自体は小学生でもできるような簡便なもので、しかも植物体を破壊することなく測定できるわけで、野外での測定や、同じ葉を経時的に測定していくような場合に力を発揮します。また、最近は、測定装置を海に沈めることにより、海洋における光合成を連続的にモニターするようなことも行なわれています。さらには、光を測定していることから、時間分解能は極めてよく、1ミリ秒以下の時間変化まで測定可能です。これを利用して、光化学系反応中心内の電子伝達反応といった極めて速い変化の解析も行なわれています。
パルス変調という手法の導入により、クロロフィル蛍光の測定は、極めて高感度に行なえるようになりました。その結果、右の写真のように、植物の葉に測定装置のグラスファイバーの先端を近づけるだけで光合成の測定が可能になりました。測定自体は小学生でもできるような簡便なもので、しかも植物体を破壊することなく測定できるわけで、野外での測定や、同じ葉を経時的に測定していくような場合に力を発揮します。また、最近は、測定装置を海に沈めることにより、海洋における光合成を連続的にモニターするようなことも行なわれています。さらには、光を測定していることから、時間分解能は極めてよく、1ミリ秒以下の時間変化まで測定可能です。これを利用して、光化学系反応中心内の電子伝達反応といった極めて速い変化の解析も行なわれています。
(2)クロロフィル蛍光画像解析
クロロフィル蛍光の測定では、その光を使うという性質から、カメラを使って2次元の情報を集めることもできます。例えば、植物の葉でも斑入りの場合は、場所によって光合成の能力が違うことが予想されますが、従来の方法では、それをきちんと調べることができませんでした。しかし、クロロフィル蛍光をCCDカメラなどで2次元的に測定して画像処理すれば、光合成能力を2次元画像として捉えることができます。
この画像解析をいろいろな植物の光合成の解析に利用した例は、「光合成実験室」に載せてありますので、ご覧頂ければと思います。いわばカメラで撮るだけなので、スイカやキウイの実などといった普通は光合成を測りづらいものでも、簡単に測定ができるのが特徴です。
(3)リモートセンシング
上記のような光合成測定とは別に、クロロフィル蛍光を単にクロロフィル量の指標として使う場合もあります。この方法が、特に有効なのは、地球規模のリモートセンシングの場合で、海洋における植物プランクトンの量を地球規模で見積もることにより、地球全体の光合成による物質生産に関する情報を知ることができます。
おわりに
ここで紹介した、光合成の測定方法は、研究の現場で使われる装置を用いたものが主で、残念ながら、教育現場に取り入れるには、ややハードルが高いかと思います。しかし、近年、大学などでも、一般向けの啓蒙活動に力を入れる場合が多くなり、筆者も、オープンキャンパスの際に、パルス変調や、蛍光画像解析を用いた測定を、一般の方々に体験して頂きました。その際の結果は「光合成実験室」に公開してあります。今後も、面白い測定のアイデアなどありましたら、試していきたいと思いますので、ご意見などございましたら、「光合成質問箱」にお寄せ下さい。
(2005年8月10日執筆、初出:高校理科研究 11, 2-7、大日本図書、2005年)
注:ヨウ素デンプン反応の項の中で、デンプンを貯めない植物の例としてホウレンソウを挙げていましたが、これは八百屋さんで買ってきたホウレンソウの場合の話で、栽培されているホウレンソウをそのまま日中に採取して調べればデンプンを持っています。誤解を招くことから、「イネ科の植物など」という記述に修正しました。(2011.12.2)