光環境変化に応答して光合成を調節する因子の解析
樋口美栄子
光合成のエネルギー源である光は植物にとって最も重要なものであり,最適な光環境を保つため植物は様々な工夫をこらしています.特に過剰な光エネルギーは活性酸素種を生じさせ植物体に障害を引き起こすため,植物はこのような障害から身を守る応答機構を発達させてきました.例えば,光合成系は強光に長時間さらされると,様々な調節によって光の吸収量を減らし,また炭酸固定系や活性酸素消去系を活発化させて新たな光環境に適応できる状態になることが知られています.このように強光応答時の生理学的な研究は古くから行われてきましたが,その応答を誘導し光合成系を調節する一連の因子についてはほとんどが未解明です.環境変化を伝える最初のシグナルをとってみても,フィトクロムなどの光受容体や光合成電子伝達鎖の酸化還元状態などが働いている可能性が示唆されていますが,一致した見解は未だ得られていません.
私は,強光応答に欠損を持つ変異体を単離することを目的としてスクリーニングを行いました.材料としては,ゲノムの全塩基配列が決定されたモデル生物であるシロイヌナズナにランダムに変異を導入したT-DNAタグラインを用いました.またスクリーニングの手法には,クロロフィル蛍光を二次元画像として経時的に測定できるシステムを用いました.クロロフィル蛍光は光合成系の状態をよく反映することが知られており,古くから光合成研究において用いられています.これまでに,クロロフィル蛍光挙動が野生株と異なる候補株を25個体単離しました.現在は,これらの候補株の詳細な特徴づけを行っているところです.単離した変異体を解析することにより,植物の環境応答,特に強光時に光合成を調節する因子およびシグナル伝達系を明らかにすることを目的として研究を進めています.
以下は修士課程の時に行った仕事です。この結果は、既に
Higuchi, M., Noguchi, T. and Sonoike, K. (2003) Over-reduced states of the Mn-cluster in cucumber leaves
induced by dark-chilling treatment. Biochim. Biophys. Acta, 1604, 151-158.
として公表されています。
低温感受性植物を用いた光合成の酸素発生系の失活と構築過程の解析
樋口 美栄子
序論
植物が行う光合成活動の一連の反応は,葉緑体のチラコイド膜に存在する電子伝達鎖で行われる.このうち光化学系II複合体上の4つのマンガン原子が配位したクラスター(マンガンクラスター)において,酸素発生反応が行われる.マンガンクラスターは,光に依存した反応でマンガンイオンからクラスター状に組み立てられることが知られており,この構築過程は光活性化と呼ばれている.
光活性化の研究は,従来 in vitroにおいて多く行われてきた.例えば,光化学系II標品をヒドロキシルアミンなどの還元剤で処理すると,光化学系IIからマンガンが抽出され酸素発生活性が失われる.しかし,ここにマンガンイオンを添加し光を照射することにより活性は回復することが知られている.光活性化の研究はこのような酸素発生の回復過程を観察することによって進められてきたが,マンガンクラスターの構築過程については未だ明確な結論には至っていない.
また,in vivoにおける光活性化の報告例は少なく,in vitroの結果が生体内でも反映されるかどうかは不明であり,in vivoにおける実験系の構築が望まれている.
低温感受性植物であるキュウリの葉を暗所で氷上におくと,酸素発生活性が大きく減少する.しかし,この活性の減少は葉を室温に戻し弱光を照射することにより回復することが報告されている.私はこのキュウリの葉を低温・暗処理するという条件がin vivoにおける光活性化のよいモデル系になりうると考えた. そこで,本研究では酸素発生系の構築過程の解析を目的として,キュウリ葉の低温・暗ストレスによる酸素発生系の阻害と光による回復過程を解析した.
結果と考察
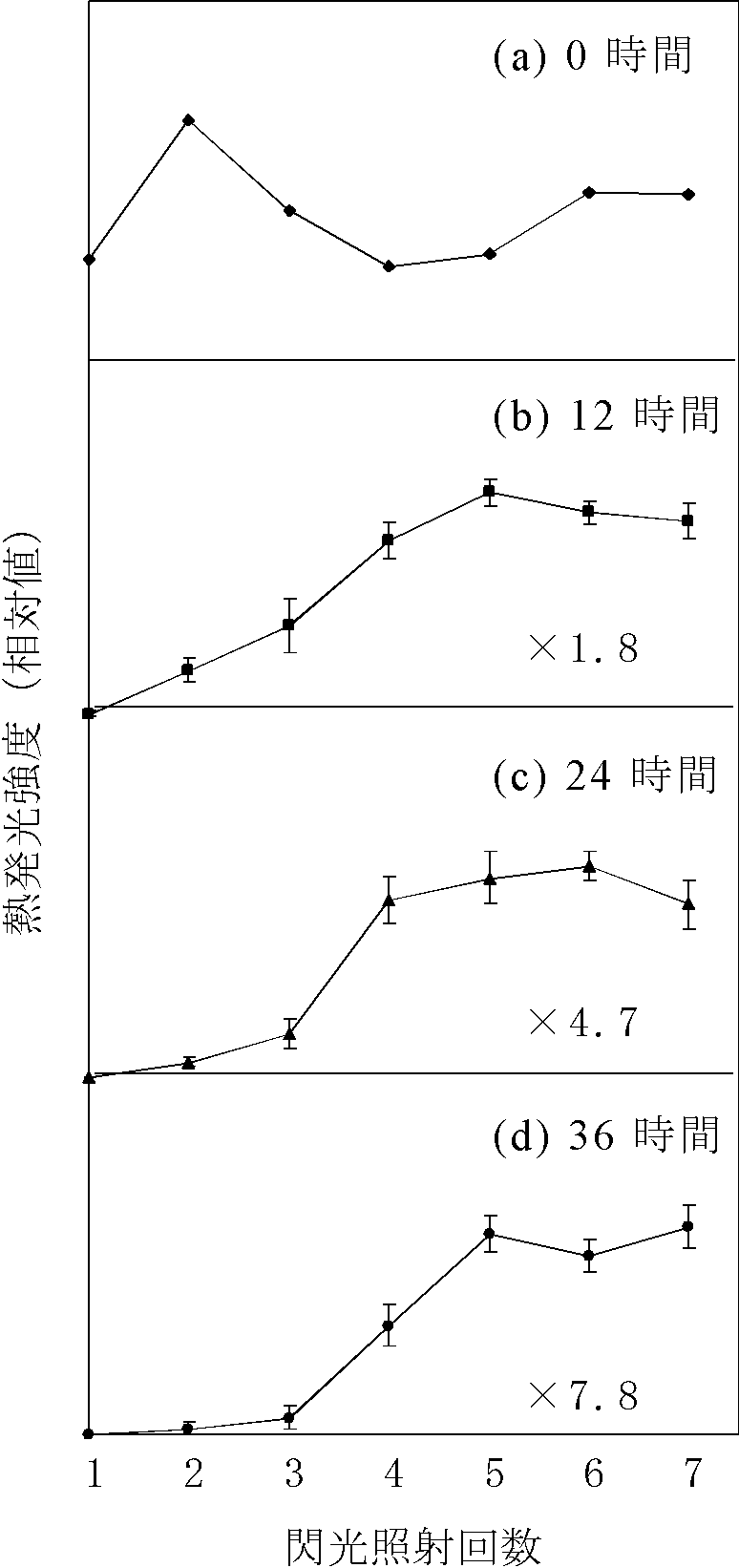 1.キュウリ葉の酸素発生系は低温・暗ストレスにより阻害される:
まず,低温・暗処理したキュウリ葉の酸素発生活性を経時的に観察したところ,活性は12時間以降徐々に低下した.
次に,低温・暗処理が酸素発生系のマンガンクラスターに与える影響を調べるため熱発光測定を行い,酸素発生活性を示す熱発光シグナル(30℃でピークを示すカーブ)を指標としてマンガンクラスターを観察した.未処理の葉で熱発光測定を行うと,1回の閃光照射で熱発光シグナルが得られる.しかし,それぞれ12,24,36時間低温・暗処理した葉で熱発光を測定したところ,処理時間が長くなるにつれ熱発光シグナルの出現に2,4,5回とより多くの閃光照射を必要とすることがわかった(右上図).この結果から,マンガンクラスターは通常のS状態(S0-S4)よりも還元された状態(S-1,S-2,S-3)として存在することが示された(右下図).低温・暗処理した葉から光化学系II標本を単離し,原子吸光光度計を用いて光化学系IIに結合しているマンガン量を測定した.その結果,マンガンは処理時間の増加に伴い減少していた.この結果は,光化学系IIからマンガンクラスターが解離したことを示している.このように還元されたマンガンクラスターとマンガンの解離は同時に観察されることから,
低温・暗処理によりマンガンクラスターが還元され,その結果マンガンイオンとして光化学系IIから解離し,酸素発生活性が減少したことが考えられる.
1.キュウリ葉の酸素発生系は低温・暗ストレスにより阻害される:
まず,低温・暗処理したキュウリ葉の酸素発生活性を経時的に観察したところ,活性は12時間以降徐々に低下した.
次に,低温・暗処理が酸素発生系のマンガンクラスターに与える影響を調べるため熱発光測定を行い,酸素発生活性を示す熱発光シグナル(30℃でピークを示すカーブ)を指標としてマンガンクラスターを観察した.未処理の葉で熱発光測定を行うと,1回の閃光照射で熱発光シグナルが得られる.しかし,それぞれ12,24,36時間低温・暗処理した葉で熱発光を測定したところ,処理時間が長くなるにつれ熱発光シグナルの出現に2,4,5回とより多くの閃光照射を必要とすることがわかった(右上図).この結果から,マンガンクラスターは通常のS状態(S0-S4)よりも還元された状態(S-1,S-2,S-3)として存在することが示された(右下図).低温・暗処理した葉から光化学系II標本を単離し,原子吸光光度計を用いて光化学系IIに結合しているマンガン量を測定した.その結果,マンガンは処理時間の増加に伴い減少していた.この結果は,光化学系IIからマンガンクラスターが解離したことを示している.このように還元されたマンガンクラスターとマンガンの解離は同時に観察されることから,
低温・暗処理によりマンガンクラスターが還元され,その結果マンガンイオンとして光化学系IIから解離し,酸素発生活性が減少したことが考えられる.
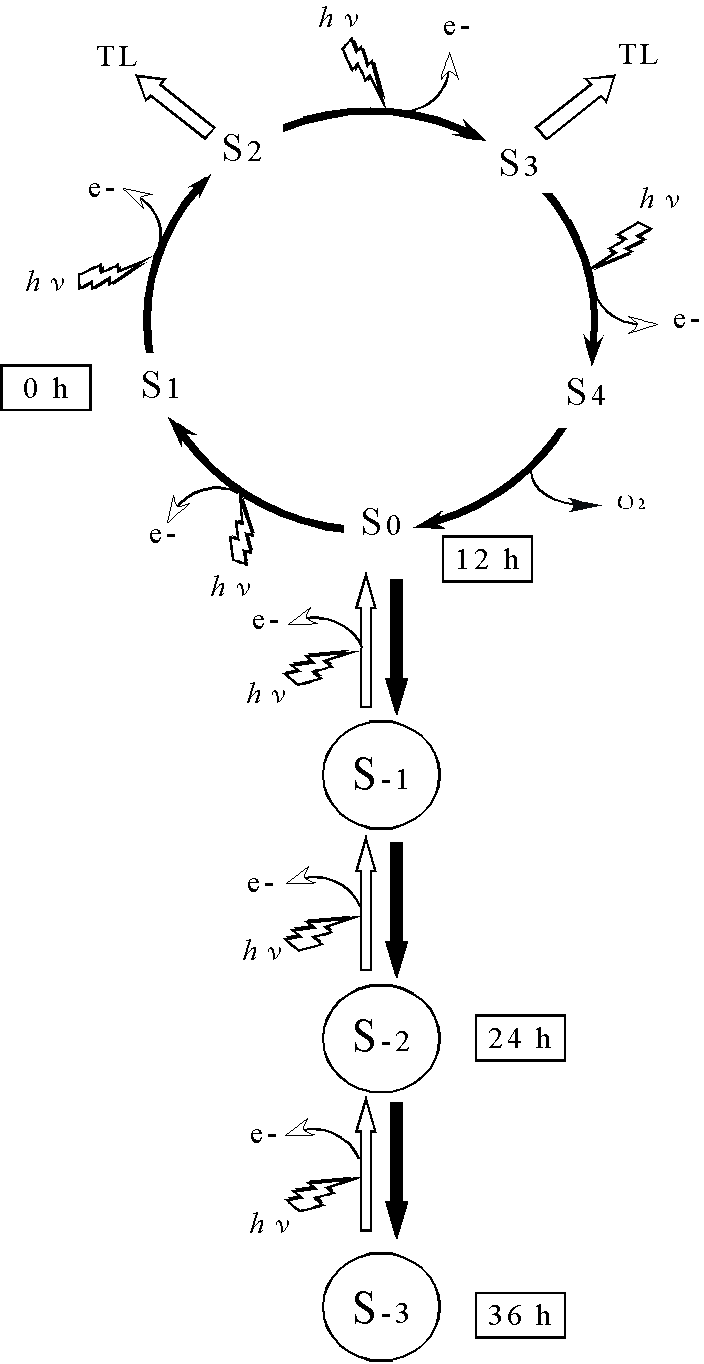 2.酸素発生系は光照射により回復する(光活性化):24時間低温・暗処理した葉を室温に戻し弱光を照射したところ,酸素発生活性は80%まで回復した.それと同時に光化学系IIに結合するマンガン数も増加した.これらの結果から,光照射によりマンガンクラスターが再構成され,酸素発生が回復したことが示された(光活性化).この光による回復過程を詳細に調べるため,閃光照射による活性化の効率を熱発光測定により観察した.その結果,熱発光シグナルの増加は二相性を示すことがわかった.熱発光強度は4回の閃光照射で約30%まで上昇し,その後は徐々に増加し500回の閃光照射で飽和に達した.この結果から,還元状態のマンガンクラスターは数回の閃光照射により酸化され活性な状態となり,解離したマンガンは500回の閃光照射により再構築されることが示唆された.
2.酸素発生系は光照射により回復する(光活性化):24時間低温・暗処理した葉を室温に戻し弱光を照射したところ,酸素発生活性は80%まで回復した.それと同時に光化学系IIに結合するマンガン数も増加した.これらの結果から,光照射によりマンガンクラスターが再構成され,酸素発生が回復したことが示された(光活性化).この光による回復過程を詳細に調べるため,閃光照射による活性化の効率を熱発光測定により観察した.その結果,熱発光シグナルの増加は二相性を示すことがわかった.熱発光強度は4回の閃光照射で約30%まで上昇し,その後は徐々に増加し500回の閃光照射で飽和に達した.この結果から,還元状態のマンガンクラスターは数回の閃光照射により酸化され活性な状態となり,解離したマンガンは500回の閃光照射により再構築されることが示唆された.
3.酸素発生系の阻害は光化学系IIの構成成分に影響を及ぼす:次に,低温・暗処理した葉から単離したチラコイド膜を用いて熱発光測定を行ったところ,5℃にピークを持つ熱発光シグナルが現れた.このシグナルは光化学系IIの電子伝達のうち,QAからQBの電子伝達が妨げられた時に観察される.このことから,低温・暗処理によりチラコイド膜のQB領域になんらかの障害が生じていることが示唆された.低温・暗処理した葉を室温に戻し弱光を照射すると,このシグナルは観察されなくなった.この結果から阻害されていた電子伝達が光照射に伴って回復することが示唆された.
また,低温・暗処理により光化学系IIの表在性タンパク質であるPsbP,PsbQタンパク質が解離することが報告されている.そこでこれらの表在性タンパク質の回復過程における挙動をSDS-PAGEで観察した結果,2つのタンパク質は室温で光照射した時に光化学系IIに再結合することが観察された.つまり,光活性化に伴ってPsbP,PsbQタンパク質は光化学系IIに結合することが示された.
以上の結果から,キュウリの葉を低温・暗処理することによりマンガンクラスターが還元され,その結果光化学系IIから解離し酸素発生活性を失うことが示された.それと同時にPsbP,PsbQタンパク質が光化学系IIから解離し,光化学系II内の電子伝達も阻害されることが明らかになった.しかし,この阻害は光照射により次のように回復すると考えられる.マンガンは光活性化により光化学系IIに還元された状態(S-3,S-2,S-1)として結合し,さらに酸化されて酸素発生活性を示す状態となる.マンガンクラスターの光活性化と同時に,PsbP,PsbQタンパク質は光化学系IIに結合し,光化学系II内の電子伝達も回復する.本研究で観察されたマンガンの酸化還元状態の変化は未だ謎の多い光合成の酸素発生反応の理解に貢献できると考えている.