オンライン教科書
パルス変調とクロロフィル蛍光を用いた光合成の測定ー測定編ー
Last revised on July 9, 2003
著作権者:園池公毅
パルス変調の測定が可能な測定機器は、現在さまざまな会社から発売されています。ここでは、実際の測定にあたって注意する点を考えていきたいと思います。このような細かい点は、どうしても機種依存になりますが、なるべく応用が利くような表現にするようにつとめました。ただし、記述は、筆者が実際に使っている機種についてのものになりがちです。この点はご容赦下さい。現在、まだ未完成ですが、一部でも皆様の参考になるかと思い、アップロードすることにしました。2003年のお正月に2.1を1項目だけ足しました。また、3月に2.2を、7月に2.3を足しました。
なお、この教科書は、WEBで公開する蛍光と光合成の教科書ですが、著作権は放棄していません。自由に、ご覧になり、プリントアウトしても構いませんが、個人としての利用に限ります。個人以外の利用は園池までご相談下さい。また、随時、修正・加筆を行いますので、この教科書を人に教える際は、プリントアウトを渡すのではなく、アドレスを教えるようにしてください。アドレスは http://www.photosynthesis.jp/fluo2.html です。なお、理論編が http://www.photosynthesis.jp/fluo1.html に出ています。
目次
1.蛍光の測定条件
1)測定光の設定
2)飽和パルス光の設定
3)励起光の設定
4)その他の設定項目 感度、応答速度
2.蛍光測定のさまざまな手法
1)基本的なクエンチング測定
2)qNの分析
3)サイクリックな電子伝達
4)QAの光還元速度
5)QAの再酸化速度
3.吸収測定の手法
1)P-700の定量
2)光化学系間のプールサイズの決定
4.さまざまな測定装置
1)基本的なパルス変調蛍光測定装置
2)高感度蛍光測定装置
3)キセノンPAM
4)二重変調蛍光測定装置
5)蛍光画像解析装置
1.蛍光の測定条件
1)測定光の設定
パルス変調における測定光は、通常1μ秒程度の長さのパルス光です。WalzのPAMシステムでは、650 nm付近に発光極大があるLEDが測定光に使われ、周波数、強度を最大に設定したときの平均光量子密度は10 μmol m-2 s-1程度です。パルス変調による蛍光測定の眼目の1つは、測定光による光合成の励起がほとんどなくすことができる点です。しかし、設定がいい加減では、これが実現しない場合があります。光合成系が励起されているかどうかは、測定パルス光をonにしたのちに、蛍光強度が変化していないかどうかでチェックできます。測定パルス光はきわめて短い時間(WalzのPAMシステムの場合100 μ秒以下)で安定するので、もし、スイッチを入れたあとに蛍光強度の増加が見られたら、測定パルス光による励起効果と考えられます。測定パルス光の強度を高く設定しているときは、測定の直前まで測定パルスをoffにしておくようにするのも1つの手です。
測定パルス光の強さは、1)パルスの持続時間、2)パルスの強さ、3)パルスの周波数、という3つのパラメーターのかけ算で決まります。このうち、パルスの持続時間は変えられません。また、パルスの周波数も、通常固定か、2種類の切り替えであり、連続的には変えられません。従って、実質上は、測定パルスの強さを変化させて、励起効果のない最大限の強さで測定すればよいことになります。
測定パルスの周波数が切り替えられる場合は、通常、低い周波数で測定を開始し、連続励起光の照射開始時、もしくは、励起用フラッシュ光の照射の直前に、高い周波数に上げることになります。WalzのPAMではこの切り替えをAuto 100 kHzモードという方法で自動的に行えます。この場合、トリガーによってフラッシュ光照射の3ミリ秒前に周波数を切り替えることもできるので便利です。測定パルスの周波数が高いと、励起効果も高くなりますので、最初から周波数を高くしておくと、正確なFoの測定に支障が出かねません。しかし、一方で、周波数が高い方が、より大きなシグナルが得られ、従ってノイズ(S/N比)を減らすことができます。また、1 kHzの周波数で測定すれば、測定間隔は1秒に1000回なので、1ミリ秒より速い変化は検出できませんが、100 kHzの周波数で測定すれば、測定間隔が1秒に10万回なので、10マイクロ秒の変化まで捉えることができるようになります。従って、QAの酸化還元のような速い時間スケールの測定を行う場合は、高い周波数での測定が必須となります。励起光があたっているときは、通常、励起光の方が測定パルス光(たとえ高い周波数でも)よりも充分強いので、測定パルス光を高い周波数にしても測定には影響を与えません。そこで、励起光を与える直前に周波数を切り替えるわけです。ただ、QAの測定の時のように単発のパルスの直前に高い周波数に切り替える場合は、パルスのあと、励起光がない状態で高い周波数での測定を続けると、蛍光強度変化の遅い成分が強調された形になる可能性があることを頭に置いておく必要があります。このような場合は、タイマーを使って10ミリ秒程度でもとの周波数に戻すのがよいでしょう。
2)飽和パルス光の設定
飽和パルス光はQAを完全還元するための光です。飽和パルス光はQAを完全還元するのに充分なだけ強くなくてはならない一方、光合成系への一般的な影響を最小限にするため必要以上に強くてはいけません。飽和パルス光の強さも、パルスの持続時間と強度のかけ算で決まります。一般的に、高等植物の葉では、連続光源からの800ミリ秒程度の持続時間に設定したパルスが使われます。WalzのPAMシステムで使われる連続励起光光源FL103はSchottのKL1500という光源を改造したものですが、スイッチをオンにした直後の過電流を避けるため、最初の300ミリ秒は光が照射されず、さらに最大の光強度に達するまでに200ミリ秒かかります。従って、持続時間を800ミリ秒に設定した場合、実際に所定の光強度があたっている時間は、800-300-200=300で300ミリ秒ということになります。この点に注意する必要があります。パルス強度は、実際に何段階か変えてみて、一番高いFmを与える強度を選ぶのがよいでしょう。 変調がうまく働いているときは、原理的には飽和パルス光自体は測定に影響を与えませんが、飽和パルス光があまりにも強いと飽和パルスの反射光などが測定フォトダイオードに入ることによる影響を充分に除けなくなります。その場合は、光源に700 nm以下の光だけを通すshort-passフィルターを入れることにより、測定フォトダイオードを保護することができます。ちなみに、WalzのPAMシステムでは、測定光の平均光量子密度の100万倍までの非変調光は、変調作用によりシステムに影響を与えないようになっています。
連続光源を使う代わりに、閃光を飽和パルス光として使う場合もあります。閃光の場合、持続時間は50ミリ秒程度必要となります。この場合、持続時間は短いので、強度は最大にしても大丈夫でしょう。ただし、50ミリ秒という短い持続時間では、記録計のペンの速度が追いつかない場合がほとんどなので、Walzのマルチプルターンオーバーフラッシュについているようなデータ保持機能を通して記録計に出力するか、オシロスコープやパソコンへデータを取り込んで処理する必要があります。なお、高等植物とは異なって、シアノバクテリアや藻類では、非光化学消光による蛍光強度の減少が非常に速い時間スケールで起こります。この場合も、正確なFmの測定のためには、データ保持機能やオシロスコープ、パソコンの使用が必要となります。飽和パルス光源としてWalzのマルチプルターンオーバーフラッシュを使うときには、光が飽和しているかどうかについては確認する必要があります。僕の経験では、FL103(KL1500)を使った場合に比べて蛍光収率が低いので、完全には飽和していないように思います。問題がないときには、FL103を使った方がよいかと思います。
3)励起光の設定
励起光は光合成系を一定の速度で駆動するための光です。従って、励起光をどれだけの強さにするのかは、実験の目的によって異なります。一番よいのは、励起光の強さをふって、励起光強度依存性をきちんと測定することでしょう。シアノバクテリアにおけるqNの励起光強度依存性のように、それによって新しい情報が得られる場合もあります(理論編5.9参照)。もし、1つの励起光強度に決める必要がある場合には、測定する植物または細胞を栽培・培養していた光強度で行うのがもっとも一般的です。いずれにせよ、蛍光の各パラメーターは励起光の強度に依存して変化しますので、実験条件としてきちんと論文に明記することが必要です。
4)その他の設定項目 感度、応答速度
感度(ゲイン)は、信号の増幅度です。当然高ければ、シグナルは大きくなりますが、一方、ノイズも大きくなります。従って、きちんとシグナルが見えている範囲内で低い方がよい、と言えるでしょう。シグナルの大きさは、前述のように測定パルスの強度にも依存しますから、もし、ノイズが非常に大きく、かつ、Foの正確な決定が目的でない場合(例えばφIIの情報だけ必要な場合など)は、感度を上げる代わりに測定パルスの強度を上げるという方法もあります。いずれにせよ、感度と測定パルスの強度の組み合わせで考える必要があります。
応答速度(レスポンス、ダンピング)は、測定回路が、どれだけ速い変動にまで追随するかを決めます。応答速度が速ければ(ダンピングが小さければ)、速い変化もきちんと捉えることができますが、応答速度が遅いほうが信号が平均化されますので、ノイズが小さくなります。QAの酸化還元のような速い反応を測定する場合は、応答速度(レスポンス)を最大にする(ダンピングを最小にする)必要があります。連続励起光による速い蛍光誘導期現象の測定では1ミリ秒程度の応答速度、秒単位の変化だけを測るのだったら、0.1秒程度の応答速度で充分です。基本的に、測定したい変化の内、最も速い変化の速度定数より、少し速い応答速度を選ぶことになります。Foの測定は、基本的に測定値が変化しません。従って、一番遅い応答速度にしても構わないことになります。DCMUの存在下でQAからQBへの電子伝達が阻害されている場合や、低温での測定では、QAの再酸化速度が遅くなるので、測定光の励起効果を最低限にしないと正確にFoが測定できません。 測定光強度を下げるとどうしてもノイズが大きくなりますが、ここで、応答速度を一番遅くすることにより、ノイズを抑えることができます。
このほか、測定系にファイバーオプティクスを使った系では、光源や測定フォトダイオードと、ファイバーオプティクスの先端の距離を変えることにより、測定光・励起光の強度やシグナルの出力を微調整することができます。これは、出力を微調整できないタイプの光源を使っているときに便利なテクニックです。
2.蛍光測定のさまざまな手法
1)基本的なクエンチング測定
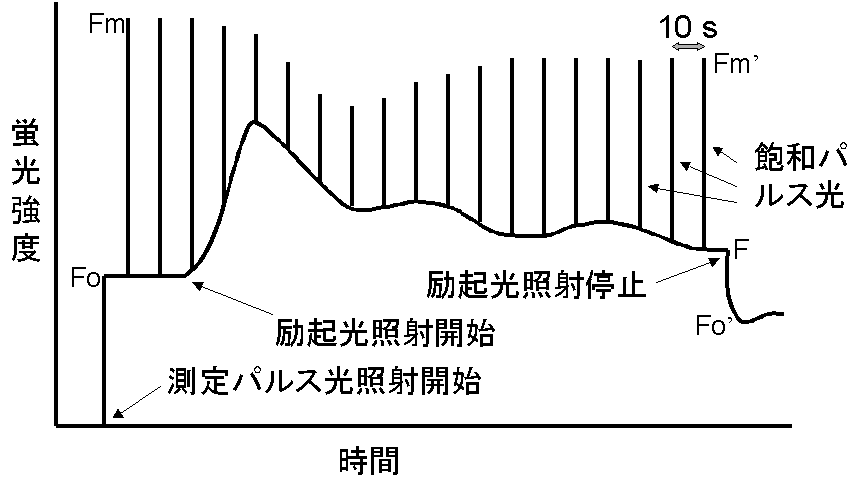 右の図は、理論編にも載せた基本的な蛍光クエンチング測定の際の測定手順です。この測定によって得られるそれぞれの時点における蛍光の強度(Fo,
Fm, Fm', F, Fo')から蛍光パラメータを計算することにより光合成の状態を知ることができます。パラメーターの意味及び計算方法は理論編を見ていただくとして、ここでは、実際の測定にあたっての注意点を考えてみましょう。
右の図は、理論編にも載せた基本的な蛍光クエンチング測定の際の測定手順です。この測定によって得られるそれぞれの時点における蛍光の強度(Fo,
Fm, Fm', F, Fo')から蛍光パラメータを計算することにより光合成の状態を知ることができます。パラメーターの意味及び計算方法は理論編を見ていただくとして、ここでは、実際の測定にあたっての注意点を考えてみましょう。
まず、測定前にどれだけ暗順応したらよいのか、という問題があります。これは、測定が何を目的としているかによって異なります。もし、電子伝達の量子収率φIIのみを知りたいのでしたら、パラメーターの算出にはFmやFoは必要ないので、そもそも暗順応する必要はありません。場合によっては、励起光すら必要でなく、栽培・培養している所へ、単にグラスファイバーを近づけて測定すれば生育光強度でのφIIは求まります。次に、ストレス処理などをした後の光合成効率の変化などを知りたいときには、暗順応は5分程度で測定することになります。ストレス処理による光合成の効率(量子収率)の変化(例えば非光化学消光などによる)は、処理修了後、徐々に小さくなりますから、あまり長く暗順応してしまうと、結局処理の効果がなくなってしまいます。逆にその植物の葉の最大の能力を知りたいときには、非光化学消光などがほぼ解消すると思われる30分以上の暗順応が必要となります。 ただ、この際、冬季の樹木の葉などでは、日中の低温光ストレスによる光合成効率の低下が、夜の間に完全には回復しない例なども報告されており、30分おけば最大能力が求まるとは断言できません。他に基本的な点としては、上に書きましたように、FoやFmがきちんと測れるような測定光強度、飽和パルス光強度になっているかどうか、励起光強度としてどのような強さを選ぶか、をきちんと押さえておく必要があります。
次に問題となるのは、どこで励起光照射を停止するかという問題です。励起光の照射を開始したあと、一時的な蛍光収率の変動があり、通常は数分でその変動が落ち着く(定常状態になる)ので、そこで励起光を停止してFo'を測定します。しかし、場合によっては、だらだらと蛍光収率が(たいていは低い方へ)変化し続けるような場合もあります。特に、生育光と異なる光環境においた場合など、なかなか定常状態にならない場合があります。場合によっては、数時間蛍光強度が変化し続ける場合もあるので、基本的にはどこかであきらめて測定するしかありません。そこを準定常状態と考えるしかないと思います。また、一定値になるのを待っているとFoより蛍光が小さくなってしまう場合もあります。これは、光が非常に強い場合や強いストレス条件下などに、光阻害が時間と共に進行するために、蛍光強度がどんどん小さくなることが原因である可能性があります。この時は光合成の励起光をもう少し弱くしてみることで解消するかどうかをみます。あまりにも強いストレス条件下での測定は、どちらにせよ何を意味するかわかりませんので、お薦めしません。 ただし、シロイヌナズナの変異株などでは、強光下で必ず定常状態の蛍光強度F(FsとかFt等とも表記)がFoを下回る例が報告されています。この場合は、非光化学消光が非常に大きいためと考えられます。従って、何らかの理由で、非光化学消光が非常に大きい場合は、野生型でも同様のことが起こる可能性があります。非光化学消光が大きい場合は、励起光を切ったあと、一旦Fo'まで少し下がり、次いで、蛍光強度がFtを越えて大きく増大し、Foまで回復します。このような変化が見えているときは、非光化学消光のせいだと判断して、そのまま実験を続ければよいかと思います。
2)qNの分析
 理論編の6.6に説明しましたように、非光化学消光(qN,
NPQ)の原因は1)チラコイド膜のプロトン濃度勾配ができたときのキサントフィルサイクルなどによる過剰エネルギー消去、2)アンテナ複合体の反応中心間の移動(ステート変化)による蛍光収率の減少、3)光化学系IIの光阻害による蛍光収率の減少が主なもので、高等植物においては、プロトン濃度勾配の項が比較的大きいとされます。この3つの成分を区別して測定する方法が、Quick
and Stittによって提案されています。励起光を切ったあとの非光化学消光の解消を考えると、プロトンの濃度勾配は、ATPaseを通したプロトンの流出で数十秒の時間スケールで解消しますし、ステート変化は10分程度で起こり、光阻害の修復は通常数十分の時間スケールでおきます。従って、これらの3つの要因は、励起光を消した後のqNの低下(蛍光の上昇)の時間経過を観察すれば区別することができることになります(右図)。ここでは、励起光を消したあとも飽和パルス光を当て続け、その飽和パルス光による蛍光の変化を観察していきます。この考え方は、もともとHorton
and HagueやDemmig and Winterによって提案されたものを改良したものです。
理論編の6.6に説明しましたように、非光化学消光(qN,
NPQ)の原因は1)チラコイド膜のプロトン濃度勾配ができたときのキサントフィルサイクルなどによる過剰エネルギー消去、2)アンテナ複合体の反応中心間の移動(ステート変化)による蛍光収率の減少、3)光化学系IIの光阻害による蛍光収率の減少が主なもので、高等植物においては、プロトン濃度勾配の項が比較的大きいとされます。この3つの成分を区別して測定する方法が、Quick
and Stittによって提案されています。励起光を切ったあとの非光化学消光の解消を考えると、プロトンの濃度勾配は、ATPaseを通したプロトンの流出で数十秒の時間スケールで解消しますし、ステート変化は10分程度で起こり、光阻害の修復は通常数十分の時間スケールでおきます。従って、これらの3つの要因は、励起光を消した後のqNの低下(蛍光の上昇)の時間経過を観察すれば区別することができることになります(右図)。ここでは、励起光を消したあとも飽和パルス光を当て続け、その飽和パルス光による蛍光の変化を観察していきます。この考え方は、もともとHorton
and HagueやDemmig and Winterによって提案されたものを改良したものです。
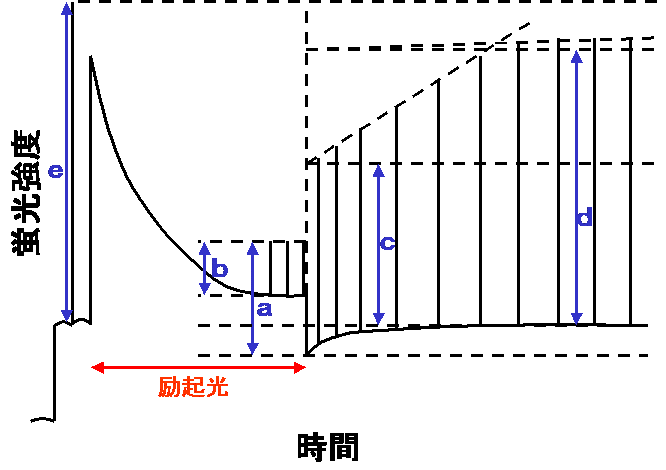
Quick and Stittの計算式では、右の図から
qf=1−a/c
qm=1−c/d
qs=1−d/e
として、蛍光消光のうち早く減衰する成分(f)、中間の成分(m)、遅い成分(s)を分けます。これらの数値は、全体の非光化学消光qNと
1−qN=(1−qf)(1−qm)(1−qs)
という関係で結びつけられます(qN=1−a/eなので計算してみるとわかります)。つまり各成分の積で全体が表されることになります。そして、それぞれの成分は、プロトン濃度勾配による項qE、ステート変化による項qT、光阻害による項qIに相当するとされます。
さて、これだけですと簡単そうで、実際にあまり考慮を払うことなく、この方法を適用している論文も見られます。しかし、実際には、3つの成分がきちんと分かれる条件は、植物によっても異なることが報告されています。考えるべき条件としては、飽和パルス光の間隔、パルス光の強さなどがあります。パルス光の間隔については、オオムギの葉では100秒以上の間隔をあければ、非光化学消光の緩和が妨害されないと報告されています。パルス光の強さについては、非光化学消光が強く誘導されている条件では、当然光捕集効率が低下していますから、より強い光が必要となります。オオムギの葉の場合は、励起光を切って5秒後の時点では、3,400
μmol/m2/sの光でもまだ飽和しないようです。励起光を切って30秒以上すれば、2,100
μmol/m2/sの光でも飽和するようなので、30秒以後のデータだけを用いれば問題ないと思われます。
このようなチェックをしさえすれば、阻害剤を用いた実験などによって、各成分がqE、qT、qIを表すことは示されていますので、プロトン濃度勾配、ステート変化、光阻害などが非光化学消光にどの程度寄与しているかを判断するには、非常によい方法だと思われます。
3)サイクリックな電子伝達
 光化学消光qPは、光化学系IIの電子受容体QAが酸化型の時に大きくなります。従って、QAが還元されると蛍光は強くなります。QAとQB、それにチラコイド膜中のプラストキノンは、分子としてはみなプラストキノンですから、タンパク質との結合によって若干変化はするものの、その酸化還元電位にはそれほど大きな違いはありません。従って、チラコイド膜中のプールのプラストキノンが還元されると、それによってQBひいてはQAが還元されることがあります。つまり、プールのプラストキノンの還元を、蛍光強度の増大として観察することが原理的には可能です。
光化学消光qPは、光化学系IIの電子受容体QAが酸化型の時に大きくなります。従って、QAが還元されると蛍光は強くなります。QAとQB、それにチラコイド膜中のプラストキノンは、分子としてはみなプラストキノンですから、タンパク質との結合によって若干変化はするものの、その酸化還元電位にはそれほど大きな違いはありません。従って、チラコイド膜中のプールのプラストキノンが還元されると、それによってQBひいてはQAが還元されることがあります。つまり、プールのプラストキノンの還元を、蛍光強度の増大として観察することが原理的には可能です。
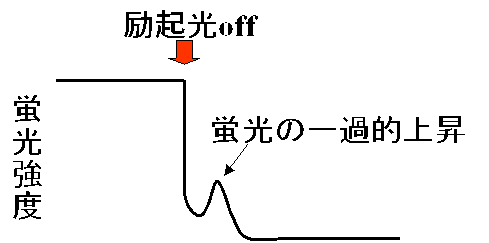
実際に、蛍光測定中に励起光を切った直後に、右図のような蛍光強度の一過的な上昇が観察されることがあります。励起光を切れば、その瞬間に光化学系の反応は停止しますから、リニアーな光合成電子伝達はほぼ停止します。しかし、励起光照射の間に光化学系Iの還元側(フェレドキシン・NADPHなど)に還元力がたまり、その還元力は光化学系Iの周りのサイクリックな電子伝達を経てプラストキノンに戻ってきます。これが蛍光の一過的な上昇を引き起こすわけです。サイクリック電子伝達には葉緑体のNDH複合体が関与していますが、奈良先端大学(現在九州大学)の鹿内先生は、この蛍光の一過的上昇を示さない変異株をスクリーニングすることで、葉緑体NDH複合体に欠損があるシロイヌナズナの変異株を得ておられます。同様な蛍光の一過的な上昇は、シアノバクテリアでも見られ、例えば、光化学系I量が多くなったような変異株では、この蛍光の上昇が大きくなることが観察されています。
サイクリックな電子伝達に関する情報は、後述するP-700の吸収変化を利用しても得ることができます。