植物の環境ストレス応答
はじめに
おそらく、多くの人にとって、「ストレス」と言えば、人間が仕事の上でなどで感じるストレスを最初に思い浮かべるのではないだろうか。そのせいか、「植物のストレス」と言うと、「植物でもストレスを感じるのでしょうか?」という質問を受けることが多い。植物がストレスを「感じる」かどうかは別として、通常の生育環境とは異なる環境におかれた植物では、その環境に応答して、さまざまな生理的変化が起こる。環境変化が激烈であれば、生育自体が阻害され、場合によっては枯死するだろうし、緩やかな環境変動に対しては、遺伝子発現や酵素反応などを制御することによって、その環境変動を乗り切ろうとする。植物にとって環境ストレスになりうる要因は、高温、低温、強光、暗黒、乾燥、降雨、塩など、さまざまであり、植物が全くストレスを受けていない理想的条件で生育できる可能性はほとんどないと言ってよい。つまり植物の生育は、どのようにストレスを乗り切るかに依存していることになり、農業的な生産性の向上や、砂漠緑化による環境改善などのためには、植物のストレス応答の理解が必須である。本稿では、植物の環境ストレス応答とそのメカニズムについて概説したい。
植物の環境ストレス応答とは
植物の環境ストレス応答はどのような特徴を持っているのであろうか。動物と植物の環境との関わりを考えた場合、一番大きな違いとなるのは動物の体の恒常性である。特に恒温動物の場合に顕著であるが、動物では外界の環境が変動しても体内の状態を常に一定に保ち、そうすることによって生存のための各種の化学反応が円滑に進行するようにしている。これに対して植物では、環境の変化は体内の状態に直接影響を及ぼす場合が多く、例えば厳寒期の樹木などにおいては、細胞自体の温度もマイナス数十℃になる。
このような動物と植物の違いは、そのストレスに対する対処法の違いにもつながる。動物では、外界の環境が変動した時に、いかに体の恒常性を保つかが重要であるが、植物では、体内の環境が変動することを前提に、そのような状況の中で、いかに生存し続けるかが重要である。このことは、植物が移動能力を持たないことによってさらに顕著となっている。植物では、強い太陽光が照りつけるからといって日陰にはいることもできず、また、雨を避けることもできない。移動能力を持っていれば避けうるストレスも、植物は甘受しなければならないのである。植物はそのような条件の中で、ストレスによる阻害を最小限に押さえ、生存に必要な化学反応を何とか進行させ続けるために、自らの代謝反応などをその状況にあったものに作りかえる。動物が体の状態を変化させないことによって生命を維持しているとすれば、植物は環境に応じて体の状態を積極的に変化さることによって生命を維持しており、そのような変化こそが、植物のストレス応答の本質であると言える。
植物にとってのストレス環境とは
それでは、植物にとってはどのような環境がストレスとなるのであろうか。一般的によく知られているのは、強光ストレス、低温ストレス、高温ストレス、乾燥ストレスなどである。この他、降雨や連続暗黒もストレスになりうる。光は、光合成により生育に必要なエネルギーを得ている植物にとって必要不可欠なものである。しかし、光エネルギーは、同時に潜在的に活性酸素の生成などを通じて光合成を阻害しうる。実際に、過剰な光の存在は強光ストレスとして光合成反応を阻害する。植物は、その生育を光合成に負っており、強光で光合成が阻害されれば当然生育も阻害されることになる。
ここで注目したいのは、その他のストレスにおける光の存在の影響である。高温ストレスの場合であれ、低温ストレスの場合であれ、単独の温度ストレスとしては致命的でないような温度でも、同時に弱い光が当たっていると、顕著な生育阻害が引き起こされる。これはその他のストレスの場合でも同様であり、暗所では生育に支障のない乾燥ストレスでも、光が当たっていると障害を引き起こす。このような相乗効果は、通常、光によって光合成反応が阻害されることによって起こっている。植物は通常光のある場所で生育しているわけであるから、その他のストレスが単独で存在することはほとんどあり得ない。実際の環境下においては、種々のストレスは光との複合ストレスである場合が多く、その場合は、光合成の阻害とその回避がストレス応答の重要なポイントになる。
環境ストレス応答のメカニズム
植物の環境ストレス応答のメカニズムは、そのストレスの種類によって大きく異なる。以下にそれぞれの場合に分けて簡単に紹介する。
強光ストレス
日の出から日没までの間に、太陽の光の強さは大きく変動する。もし、一番光が強いときにもストレスとならないような光合成の仕組みを作ったとすると、光がそれより弱い時に光合成の効率が下がってしまう。そこで、植物は、中程度の光の強さに適応した光合成系を持ち、かつ、強い光があったときにはそれに対する防御機構を発動する、という生存戦略を採用していることが多い。強光に対する防御機構としては、過剰な光エネルギーを熱エネルギーに変換して発散させる非光化学消光と呼ばれるシステムや、過剰なエネルギーによって生成した活性酸素を消去する活性酸素消去系などが知られている。これらの防御系が働かないと、光合成の2つある光エネルギー変換系のうち、光化学系IIと呼ばれる部分で活性酸素の一種である一重項酸素が生成し、これによって光化学系IIが阻害される。
温度ストレス
温度ストレスのうち高温ストレスにおいては、タンパク質の変性や複合体の解離などによる機能喪失が生育阻害の直接的な原因となると考えられる。高温ストレスに対する応答は、タンパク質の一次構造の変化による熱耐性の獲得などの進化的な適応と、熱ショックタンパク質と呼ばれるタンパク質の変性を防ぐ(もしくは変性から回復するのを助ける)タンパク質の発現などの短い時間スケールでの応答が、植物の主な対処手段となる。
低温ストレスは、freezingと呼ばれる凍結ストレスと、chillingと呼ばれる0〜10℃ぐらいの温度の低温ストレスに分けられる。freezingにおいては、細胞内で氷の結晶が成長することによる細胞の物理的な破壊と、細胞外で水が凍ることによって細胞内の水が失われ、浸透圧が上昇することによる障害が、主な生育阻害の原因となる。このため、耐凍性を持つ植物では、細胞内液をいわば不凍液化して細胞内の凍結を防ぎ、細胞内に浸透圧調節物質を貯めることによって浸透圧の上昇による障害を回避するシステムが存在する。このようなシステムは、凍らない程度の低温によって誘導されるため、徐々に温度が低下したときには、freezingに対処できるが、暖かいところから突然温度が低下したときには対処できず大きな生育阻害を受ける。
一方、chillingは単独でもストレスになりうるが、通常は光との複合ストレスを示す典型的な例である(1)。単独では、低温に弱い植物であっても、0℃付近に24〜48時間さらされて初めて阻害が引き起こされるが、太陽の直射日光の1/20程度の弱い光が伴うと、4℃付近に数時間おかれただけで光合成活性が完全に失われる。低温光ストレスによる阻害部位は、光合成の光エネルギー変換の場の1つである光化学系Iであることがわかっている(2)。chillingストレスにおいては、低温における生体膜の物性の変化が重要な役割を果たしていると考えられ、生体膜を構成する不飽和脂肪酸の変化とストレス耐性の間の相関が報告されている。
乾燥ストレスと降雨ストレス
乾燥ストレスは、細胞から水分が失われる点では、前述したfreezingストレスと同様であり、乾燥ストレスによって誘導される遺伝子群などはfreezingストレスの場合と共通のものが多い。また、乾燥ストレスの下では、水分の損失を防ぐため葉の気孔を閉じて蒸散を抑える。一方で、光合成の基質である二酸化炭素は気孔を通じて取り込まれるため、乾燥ストレスの下では、葉の中の二酸化炭素濃度が低下する。従って、乾燥ストレスは二次的に低二酸化炭素ストレスを引き起こす。興味深いことに、乾燥ストレスの対極とも思われる降雨も低二酸化炭素ストレスを引き起こす。これは、葉の表面の濡れが気孔の閉鎖をもたらすためである。降雨ストレス(3)や低二酸化炭素ストレス(4)では2つの光化学系の間をつなぐ電子伝達が阻害部位であると報告されている。
「阻害」と「調節」
上述のように環境ストレス下においては、光合成活性が低下するなどのさまざまな阻害が見られる。しかし、その阻害が必ずしも、植物にとって有害な反応であるとは限らない。以下にそのような例を見てみよう。
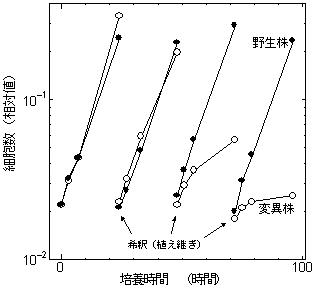 シアノバクテリアは原核単細胞生物でありながら高等植物のような光合成を行い、植物の光合成反応のモデル生物としてよく研究されている。このシアノバクテリアで、強光ストレスによる光化学系IIの光阻害が起こらないような変異株の生育を調べると、確かに24時間以内の短期的な生育は変異株が野生株を上回る。ところが、2日、3日と強光条件が続くと、やがて変異株の生育速度は低下し、やがて死滅してしまう(右図)。これは、野生株では、光化学系IIの活性を低下させることによって強光下での活性酸素の生成を抑え、強光下での持続的な生育を可能にしているためと考えられる(5)。つまり、この場合は、光合成の阻害が、光ストレスを生き延びるために必要不可欠であったことを意味する。
シアノバクテリアは原核単細胞生物でありながら高等植物のような光合成を行い、植物の光合成反応のモデル生物としてよく研究されている。このシアノバクテリアで、強光ストレスによる光化学系IIの光阻害が起こらないような変異株の生育を調べると、確かに24時間以内の短期的な生育は変異株が野生株を上回る。ところが、2日、3日と強光条件が続くと、やがて変異株の生育速度は低下し、やがて死滅してしまう(右図)。これは、野生株では、光化学系IIの活性を低下させることによって強光下での活性酸素の生成を抑え、強光下での持続的な生育を可能にしているためと考えられる(5)。つまり、この場合は、光合成の阻害が、光ストレスを生き延びるために必要不可欠であったことを意味する。
別の例を見てみよう。キュウリの葉は、chillingストレスを受けた直後でも見た目には変化がないが、常温に戻った後、3日ぐらいの間に葉の退色が見られる(下図)。このような、低温ストレス後の可視障害の進行は古くから知られてきたが、なぜ、ストレスがなくなったあとに、かえって傷害が進行するかは不明であった。最近になり、低温処理直後に光化学系Iの活性が不可逆的に失活していることが明らかとなった。活性を持たずに光のエネルギーを吸収する光化学系が存在すると、その過剰なエネルギーによって活性酸素が生じ、細胞構成成分のさらなる破壊が進む。これを避けるために、植物は、活性を失った光化学系Iをストレス後に分解し、結果としてクロロフィルが失われ葉が退色していると考えられる(6)。従って、葉の退色の進行は、さらなる傷害を避けるための、植物の積極的な方策なのである。

キュウリの葉の低温ストレス後の外観の変化: a, 未処理の葉、b, 低温処理直後、c, d, e, f, g, h, 低温処理後、室温に戻し、それぞれ、1日目、2日目、3日目、4日目、5日目、6日目。葉の約下半分が低温処理を受けている。
ストレス応答研究の応用にむけての問題点
植物のストレス応答のメカニズムの解明は、ストレス耐性植物の作出につながり、農業生産性の向上や環境改善に寄与すると期待される。しかし、メカニズムの解明が着々と進行する一方で、その応用に関してはまだまだ課題が多い。これにはいくつかの原因があると考えられる。
その中の1つの問題として、上述のように、阻害が必ずしも植物にとって害であるとは限らないということがある。前項のシアノバクテリアの例で見られたように、場合によっては、ストレスによる阻害を回避できるような植物を作り出すと、かえって生育が低下する場合さえあるかも知れない。
別の問題として、阻害部位の同定の困難さがあげられる。例えば、chillingストレスにおいては、過去に様々な条件で様々な異なる阻害部位が報告され、ストレス応答の理解に混乱をもたらしていた。これは、chillingストレスが前述のように通常複合ストレスとして現れ、ストレス時の光強度によって阻害部位が異なっていたことが、1つの原因であった。さらにストレス条件が厳しい場合には、複数の部位が阻害されるが、個々の研究によって注目する部位が異なると、報告によって阻害部位が異なることになってしまう。
chillingストレスの阻害部位は、現実の環境でのストレス条件を考え、その中でもっとも阻害を受けやすい部位を検討することによって初めて明らかとなった。1日の中で一番冷え込むのは明け方であり、その際の光条件としては弱光が想定される。そして、低温・弱光条件で阻害を受ける部位の中でもっとも感受性の高い部位を探索することによって、阻害部位が明らかとなったのである。ごく当たり前のことであるが、現実の環境での条件を無視したストレス条件における実験では応用面の成果は期待できない。
もう1つの問題としては、阻害部位の解明だけではストレス耐性の付与につながらないということがある。キュウリの葉は典型的な低温感受性を示し、弱光下のchillingによって光化学系Iが阻害される。しかし、光合成の場である葉緑体のチラコイド膜を単離して弱光を照射すると、温度によらずに、またキュウリでなく低温耐性植物のホウレンソウの場合でも、光化学系Iが阻害される(下の表を参照)。このことから、本来、光によって阻害を受けやすい光化学系Iが、葉の中では何らかの機構によって保護されており、その保護機構こそが低温感受性であると結論できる。つまり、この場合、阻害部位は光化学系Iであるが、低温感受性植物を低温感受性たらしめているのは、光化学系Iではなく、それを保護する未知のメカニズムであることになる。
| 材料 | キュウリ(低温感受性) | ホウレンソウ(低温耐性) | ||
| 処理温度 | 低温 | 室温 | 低温 | 室温 |
| 葉 | 阻害 | 正常 | 正常 | 正常 |
| チラコイド膜 | 阻害 | 阻害 | 阻害 | 阻害 |
おわりに
植物のストレス応答の研究は、ようやくその分子メカニズムに手が届き始めた段階である。前項に述べたような問題点を考えると、ストレス応答研究の成果を応用に生かすためには、単に、ストレスによる阻害部位を決定するという局所的な目的にとらわれず、植物の環境ストレス応答全体を大きなシステムとして理解することが必要不可欠であると思われる。
参考文献
筆者が直接関わった仕事についてのみ原著論文もしくは総説をあげておく。
- Sonoike, K. (1998) Various Aspects of Inhibition of Photosynthesis under Light/Chilling Stress: "Photoinhibition at Chilling Temperatures" versus "Chilling Damage in the Light". J. Plant Res. 111, 121-129.
- Sonoike, K. (1996) Photoinhibition of Photosystem I: Its Physiological Significance in the Chilling Sensitivity of Plants. Plant Cell Physiol. 37, 239-247.
- Ishibashi, M., Sonoike, K. and Watanabe, A. (1997) Photoinhibition of Photosynthesis during Rain Treatment: Identification of the Intersystem Electron-Transfer Chain as the Site of Inhibition. Plant Cell Physiol. 38, 168-172.
- Ishibashi, M., Sonoike, K. and Watanabe, A. (1997) The Inhibition of Photosynthesis after Exposure of Bean Leaves to Various Low Levels of CO2. Plant Cell Physiol. 38, 619-624.
- Sonoike, K., Hihara, Y. and Ikeuchi, M. (2001) Physiological Significance of the Regulation of Photosystem Stoichiometry upon High Light Acclimation of Synechocystis sp. PCC 6803. Plant Cell Physiol. 42, 379-384.
- Kudoh, H. and Sonoike, K. (2002) Irreversible damage to photosystem I by chilling in the light: cause of the degradation of chlorophyll after returning to normal growth temperature. Planta 215, 541-548.
(2001年10月22日執筆、初出:Ajico News 203, 1-8、2001年)